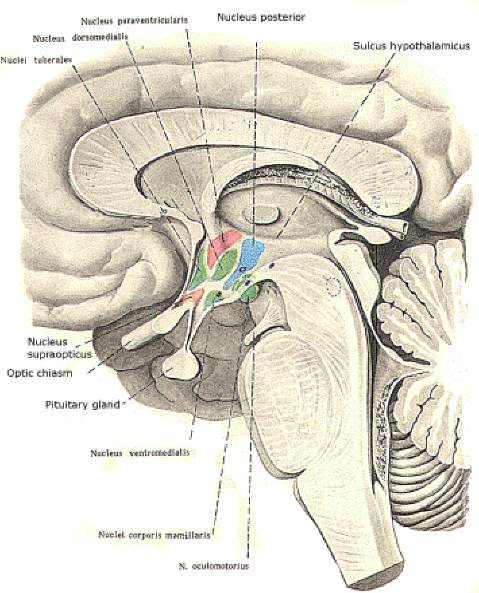
The hypothalamus forms the floor of the third ventricle and is separated from the thalamus above by the hypothalamic sulcus in the ventricle's lateral walls. It is composed of a discrete set of nuclei (Fig-1 and 2) which are involved in the following functions:
1 Autonomic control
2 Temperature regulation
3 Thirst and control of body water
4 Appetite control
5 Endocrine control
6 Emotional reactions
7 Sleep and wakefulness
8 Stress response
2 Temperature regulation
3 Thirst and control of body water
4 Appetite control
5 Endocrine control
6 Emotional reactions
7 Sleep and wakefulness
8 Stress response
Hypothalamic Nuclei
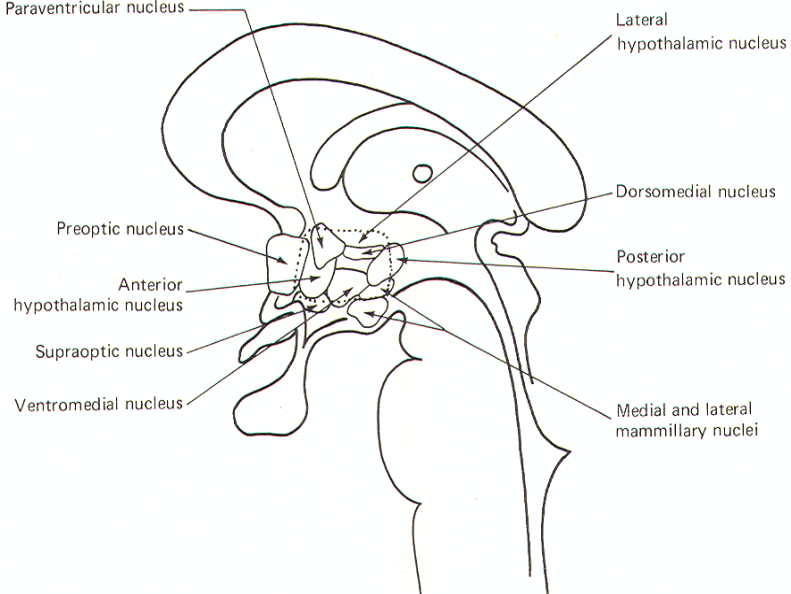
Several nuclei have been identified in the hypothalamus. Some have become associated with specific physiological activities, while the functions of others are less clear and in some cases unknown. Their relative locations are illustrated in midsagittal section in Fig-1 and 2. Therefore it is important to recognize that you are seeing the nuclei on the right side of the third ventricle only. In other words, each of the nuclei is paired. The nuclei are often grouped in four general areas. The preoptic area includes the medial and lateral preoptic nuclei, which extend through the lamina terminalis. The supraoptic area includes the supraoptic, anterior hypothalamic, and paraventricular nuclei. The tuberal area include the lateral hypothalamic, posterior hypothalamic, dorsomedial, and ventromedial nuclei. Finally, the mammillary area is composed of the medial and lateral mammillary nuclei.
 | |
| Fig-1: | Fig-2: |
For the hypothalamus to play an effective role in the functions listed above, it is necessary that it be in neural contact with many areas of the brain and spinal cord. The fiber systems involved can be described as either afferent or efferent to the hypothalamus. Some of the principal systems are presented below.
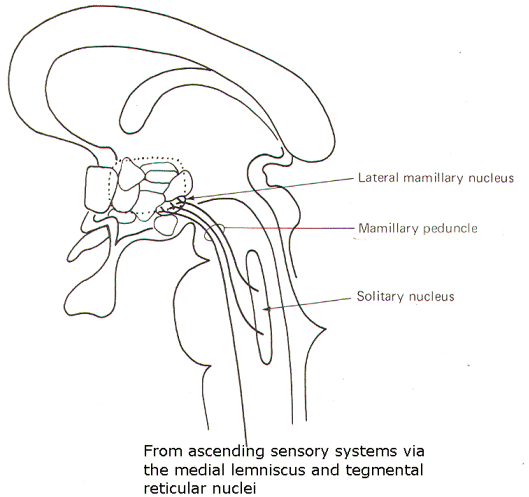
Hypothalamic Afferent Input Fibers in the mammillary peduncle represent a major ascending input to the hypothalamus (Fig-3). It arises in the tegmentum of the midbrain and is formed by fibers carrying information from SVA and GVA fibers which terminate in the solitary nucleus. Similarly, ascending information from the spinal cord relayed through the medial lemniscus also contributes fibers to this system. The hypothalamic termination is chiefly in the lateral mammillary nuclei.
  | |
| Fig-3 | Fig-4 |
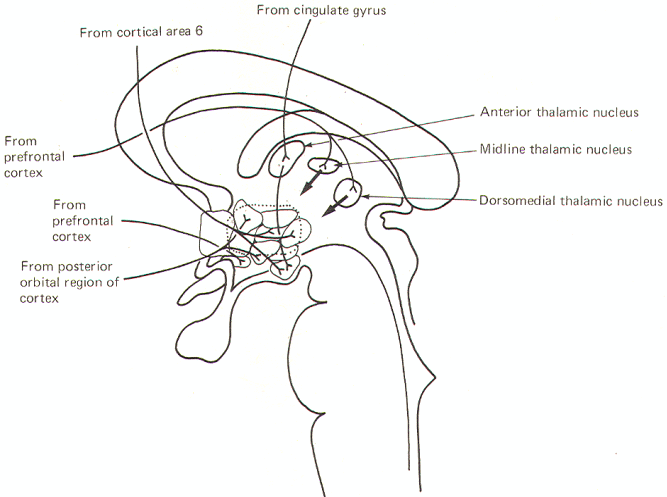
The corticohypothalamic fibers project to a number of hypothalamic nuclei. It is no doubt through such connections that conscious thought is often able to give rise to autonomic and visceral responses such as, for example, indigestion from worry, sweating from fear, and sexual arousal from certain kinds of thoughts. Nevertheless, the hypothalamus is not ordinarily under cortical control as evidenced, for example, by our inability to raise or lower the blood pressure at will.
Several corticohypothalamic routes are illustrated in Fig-4. Fibers from cortical area 6 pass through the septal region to terminate chiefly in the posterior hypothalamic and lateral hypothalamic nuclei as well as the mammillary nuclei. Fibers from the prefrontal cortex project to the supraoptic nucleus as well as indirectly to the hypothalamus through synapses in the anterior, midline, and dorsomedial thalamic nuclei. Projections from the olfactory posterior orbital region of the cortex project to the paraventricular and ventromedial nuclei. The cingulate gyrus also indirectly influences the hypothalamus via an intermediate synapse in the anterior thalamic nucleus. Thalamomammillary fibers are also present.
The thalamohypothalamic fibers fall into two general groups; the thalamomammillary fibers which project from the anterior thalamic nucleus to the medial mammillary nucleus, and a group which passes from the midline and dorsomedial thalamic nuclei principally to the anterior hypothalamic nucleus. There are probably other connections as well between the thalamus and hypothalamus (Fig-5).
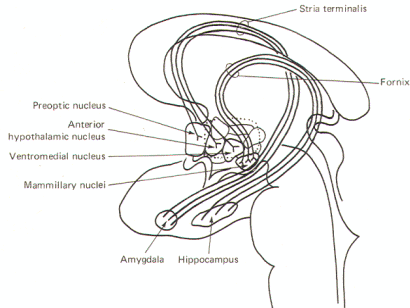 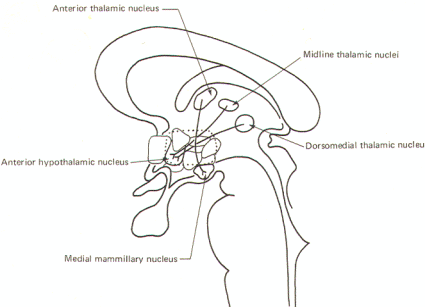 | ||
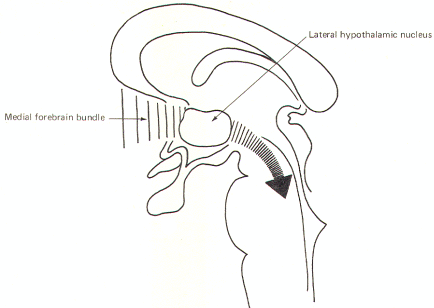 Fig-5 Fig-5 | Fig-6 | Fig-7 |
The corticomammillary fibers (fornix) project from the hippocampus of the temporal lobe to the mammillary nuclei via a long loop (Fig-6). The stria terminalis is composed of fibers which originate in the amygdala of the temporal lobe and pass caudally along the tail of the caudate nucleus and arch over the dorsal aspect of the thalamus to terminate in the septal nuclei as well as the preoptic, anterior hypothalamic, and ventromedial nuclei. The medial forebrain bundle is a complex group of fibers which arise in the basal olfactory region, the septal nuclei, and periamygdaloid region and pass to the lateral hypothalamic nuclear area (Fig-7). Many medial forebrain bundle fibers continue into the midbrain tegmentum while others project to additional hypothalamic nuclei. Those reaching the midbrain tegmentum relay signals to the autonomic and visceral controlling nuclei of the brainstem. Hence the bundle is both an afferent and efferent system with respect to hypothalamic nuclei.
Hypothalamic Efferent Output
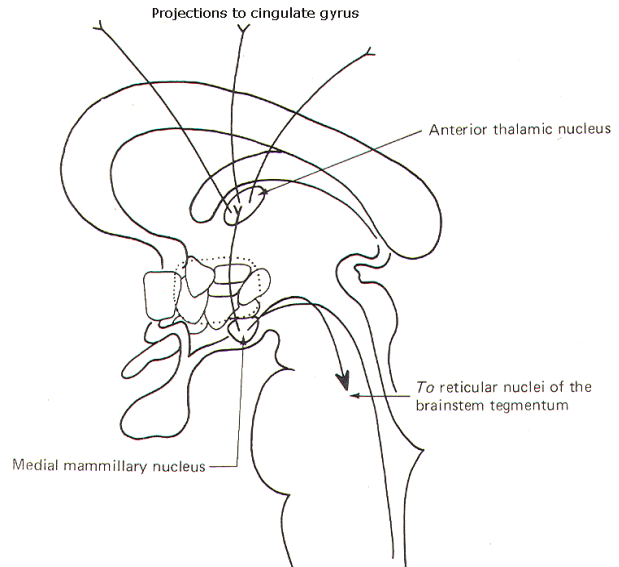
The anterior thalamic and mammillary nuclei are reciprocally related and therefore a mammillothalamic tract exists. Through projection fibers from the anterior thalamic nucleus to the cingulate gyrus, the hypothalamus is able to influence activity in this region of the cerebral cortex. This system and the mammillotegmental fibers which project to the reticular nuclei of the brain stem tegmentum are illustrated in Figure-8.
  | |
| Fig-8 | Fig-9: |
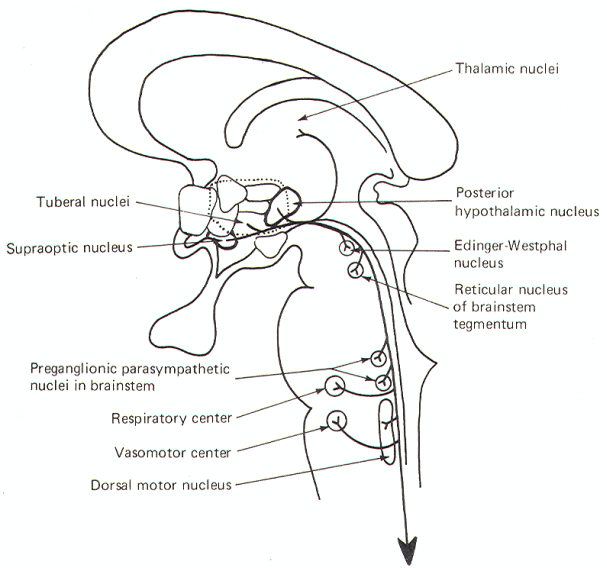
The periventricular fibers represent a large descending fiber system originating in the supraoptic, posterior hypothalamic, and tuberal nuclei. While there is a small ascending component to thalamic nuclei, most of the fibers descend to synapse in various parasympathetic brainstem nuclei as well as the respiratory and vasomotor centers. Some also terminate in the reticular nuclei of the brainstem tegmentum. Reticulospinal fibers as well as some periventricular fibers which don't synapse in the brainstem, descend into the spinal cord to influence preganglionic sympathetic and parasympathetic neurons in the intermediolateral region (Fig-9).
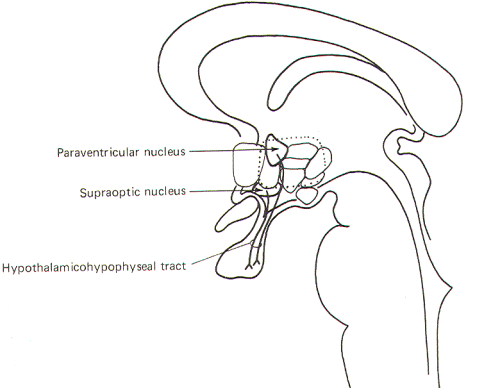
The hypothalamohypophyseal tract is a group of fibers which run from the paraventricular and supraoptic nuclei to the posterior lobe of the pituitary gland. This tract mediates release of the posterior pituitary hormones, oxytocin, and antidiuretic hormone (ADH). Oxytocin is synthesized in the paraventricular nucleus and transported through the axons of fibers projecting to the posterior lobe. ADH is synthesized in the supraoptic nucleus and similarly transported through the hypothalamohypophyseal tract to the posterior lobe (Fig-10). The hormones are stored in the terminal endings of these fibers until they are released into the circulation.
 | ||
| Fig-10 |
The Hypothalamus and the Autonomic Nervous System
The hypothalamus has long been suspected of playing a role in autonomic nervous system regulation. Most of the evidence for this is based on the observation that electrical stimulation of various areas of the hypothalamus produce autonomic effects. While there is no clear-cut demarcation line. stimulation of the caudal hypothalamus generally produces an increase in sympathetic activity, while stimulation of the rostral hypothalamus produces parasympathetic effects. It is reasonable to assume that the hypothalamus is not the sole, or even the principal, regulator of autonomic activity, While it can certainly modify autonomic activity via direct and indirect pathways to preganglionic neurons in the brain stem and spinal cord, we must also recognize that the hypothalamus itself receives input from a wide variety of sources in both the brain and spinal cord. Thus, while the hypothalamus can certainly modify autonomic response, the question of ultimate control is certainly larger and more complex than can be explained by a model based on hypothalamic control alone.
The Hypothalamus and Temperature Regulation
Temperature regulation is an important homeostatic activity which is primarily controlled by the hypothalamus. If we consider the dangerous effects of temperature extremes on the body, a center designed for regulating this variable is of obvious importance. Electrical stimulation of the anterior hypothalamus, particularly the supraoptic area, triggers a thermolytic response, That is, those activities which cause the body temperature to drop are set into operation. Conversely, stimulation of the posterior hypothalamus, particularly the tuberal area, triggers a thermogenic response, reflected both in increased heat conservation and production. Thermolytic responses include cutaneous vasodilation in order to increase heat loss by radiation, sweating to increase heat loss by evaporation, and panting in animals like the dog. Thermogenic responses include cutaneous vasoconstriction to prevent heat loss by radiation, shivering to produce heat by increased muscular activity, cessation of sweating to reduce heat loss by evaporation, and an increase in the production and release of thyroxine in order to increase the metabolic rate. Thermoreceptors in the hypothalamus are sensitive to very small changes in the temperature of circulating blood. Because blood temperature varies closely with changes in core temperature, the hypothalamus is continually kept informed of changes in the overall temperature of the body. Subsequently it can activate appropriate thermolytic or thermogenic activities in order to restore body temperature to normal. The hypothalamus also receives input from cutaneous thermoreceptors which keep it informed of changes in the environmental temperature. Consequently the hypothalamus is continually informed of both external and internal temperature changes and is well equipped through neural activation of appropriate effectors to prevent temperature fluctuations by regulating body temperature within very narrow limits.
The Hypothalamus, Thirst, and Control of Body Water
The hypothalamus is well equipped to respond to changes in the total amount of body water. A poorly localized area of the hypothalamus called the "thirst center" is stimulated by a dry mouth as well as body dehydration, Projections from the thirst center to the thalamus and then to the conscious cortex inform us of the need for water. This triggers the sensation of thirst and initiates the conscious desire for water. The hypothalamus also takes subconscious steps to correct dehydration.
Osmoreceptors in the supraoptic nuclei respond to dehydration (typically associated with increased osmolality in the circulating blood) by increasing the production and release of antidiuretic hormone (ADH). This hormone is produced in the supraoptic nucleus (SON) and transported via the axons of the hypothalamohypophyseal tract to the posterior pituitary lobe for temporary storage and ultimate release into the circulation. Once released, ADH promotes an increase in total body water by facilitating water reabsorption in the kidneys so that more is returned to the blood and less is lost in the urine. ADH operates by increasing the water permeability of the distal tubules and collecting ducts of the nephrons. This causes water to be osmotically reabsorbed from the less osmotic glomerular filtrate to the more osmotic extracellular fluid of the kidney medulla and renal blood supply.
The Hypothalamus and Appetite
Studies on animals have confirmed the relationship between the hypothalamus and appetite. The lateral hypothalamic nuclei function in part as a "feeding center." This is based primarily on the observation that electrical stimulation of this region in the rat triggers a strong feeding response which is observed even if the animal has just eaten his fill. Conversely, the ventromedial nucleus is described as the "satiety center" because stimulation of this region stops all feeding activity on the part of the animal. It is certainly possible that these two nuclei are neurally related in such a way that each inhibits the other. In this way, when the lateral hypothalamic nucleus is directing feeding, it can also simultaneously inhibit the satiety center, and vice versa. At present, the system is poorly understood in humans. If such a mutually exclusive system exists, however, it is obviously capable of conscious modification, as we can eat when full and refrain from eating even when hungry.
The Hypothalamus and the Endocrine System
If, as it is often said, the pituitary is the master gland of the endocrine system, it can equally be said that the hypothalamus is master of the pituitary. It influences the production and release of hormones from both the posterior lobe (pars nervosa or neurohypophysis) as well as from the anterior lobe (pars distalis or adenohypophysis). Unlike the anterior lobe, which is not derived from neural tissue, the posterior lobe has an intimate embryological relationship with the hypothalamus. Because of this difference, the hypothalamus exerts its influence in a different manner on each lobe.
Control of the Posterior Lobe The two known posterior pituitary hormones are oxytocin and antidiuretic hormone, also called vasopressin. Each is an ~ whose amino acid sequence is well known. There are no secretory cells in the posterior pituitary, however, and both hormones are produced in the hypothalamic nuclei and subsequently transported to the posterior lobe.
Oxytocin is probably produced in the paraventricular nucleus (PVN). Its target tissues include the breast. where it promotes the letdown of milk, and the uterine musculature. where it promotes smooth muscle contractions. It's released in response to several stimuli. These include mechanical stimulation of the nipple area by the suckling infant. uterine and cervical contractions associated with birth. and psychic factors via poorly understood circuits from the conscious cortex. The latter is apparent when the cry of a hungry infant is often a sufficient stimulus for milk letdown in the lactating mother. requiring no mechanical stimulation at all.
Antidiuretic hormone is produced in the supraoptic nucleus and similarly transported to the posterior lobe. The stimulus for its release (stimulation of the thirst center, dehydration, and increased body fluid osmolality) have previously been discussed. ADH is also called vasopressin because of its ability to va soconstrict blood vessels. Once synthesized, the hormones are transported to the posterior lobe via axonal transport through fibers of the hypothalamohypophyseal tract. Here they are temporarily stored bound to a protein (neurophysin) until their release is called for.
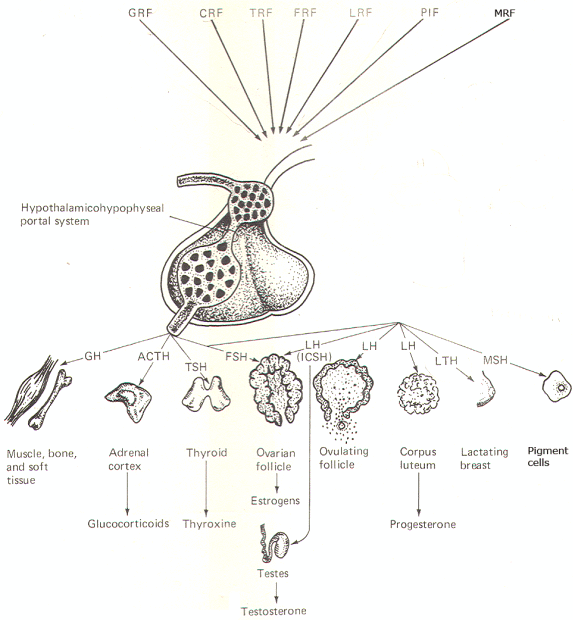
Control of the Anterior Lobe There are no direct nerve fiber pathways from the hypothalamus to the anterior lobe. And unlike the posterior lobe. it is rich in secretory cells. Thus, the hormones of the anterior lobe are both produced in and released from the adenohypophysis. The known hormones from the anterior lobe include: growth hormone (G H), adrenocorticotrophic hormone (ACTH), thyroid-stimulating hormone (TSH). follicle-stimulating hormone (FSH), luteinizing hormone (LH), luteotropic hormone (L TH), and melanocyte-stimulating hormone (MSH). Luteinizing hormone is called interstitial cell-stimulating hormone (lCSH) in the male.
While these hormones are actually synthesized in the anterior lobe of the pituitary. the signal for their release comes from the hypothalamus in the form of small polypeptides called releasing factors. At the appropriate time a particular releasing factor is secreted near the capillary network in the median eminence (Fig-11) by fibers from one or more of the hypothalamic nuclei. It then diffuses into the capillaries and travels into the adenohypophysis via thehypothalamohypophyseal portal system. Once in the anterior lobe. the portal system again gives rise to a capillary network. The releasing factor then diffuses out of the capillaries and causes specific groups of secretory cells to release their hormone into the capillaries for distribution to the main circulation. Figure 15-10 illustrates the various known releasing factors as well as their hormones and target tissues.
 | ||
| Fig-11 |
The Hypothalamus and Emotion: The Limbic System
In addition to its other functions, the hypothalamus also plays a role in the physical expression of emotion. Parts of the hypothalamus are closely integrated with the limbic lobe of the brain. This lobe. illustrated in Fig-12, includes the cingulate gyrus, isthmus, and parahippocampal gyrus and uncus. The limbic lobe together with the amygdala, hippocampus, olfactory bulbs and trigone, fornix, and mammillary bodies comprise the limbic system. In lower vertebrates this system is primarily involved with smell. However in humans, its principal role appears to be in the arousal of emotion.
The cerebral cortex is associated with the subjective aspects of "feelings" and emotions while the autonomic nervous system promotes many of the physical expressions associated with them. It does this through changes in such activities as heart rate, blood pressure, sweating, salivation. and gastrointestinal activity. One theory is that the limbic system ties the cerebral and autonomic components of emotion together. We all know that it is possible to worry enough about something to the point where it brings on physical symptoms such as stomach upset, sweating, etc.
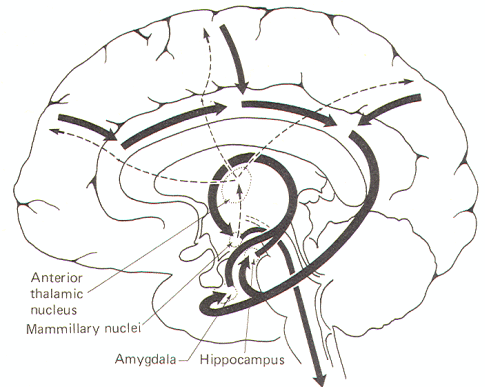
Figure-12 illustrates a model for this phenomenon. The conscious neocortex is reciprocally connected to the cingulate gyrus. which in turn transmits to the parahippocampal gyrus and uncus of the temporal lobe via the isthmus. These cortical areas project to the subcortical hippocampal and amygdaloid nuclei. Fibers projecting from these nuclei pass through the looping arch of the fornix to the mammillary nuclei. These, together with other hypothalamic nuclei. promote autonomic responses through descending fibers to autonomic nuclei within the brain stem and spinal cord.
 | ||
| Fig-12 |
The system probably works in reverse also. If strong autonomic activity is going on at a subconscious level, the conscious cortex often becomes aware of it. This awareness is probably mediated over mammillothalamic fibers which project to the anterior nucleus of the thalamus, which then project to the cingulate gyrus and the conscious cortex. It must be understood that the pathways described here certainly do not represent the complete network between the cerebral and autonomic components of emotion. This is clearly an area about which we know very little.
Hiç yorum yok:
Yorum Gönder